回顾固有免疫核心概念的发展
“免疫”几乎是所有生物体所共有的,在过去的几十年里,人们对适应性免疫有了越来越多的认识,而固有免疫在很大程度上被人们所忽视。大量工作表明固有免疫在维持体内平衡、塑造微生物群以及癌症、神经退行性疾病、代谢综合征和衰老等中都发挥着关键作用,这些功能的鉴定也伴随着相关术语及概念的出现,如“模式识别受体(PRR)”、“病原体相关分子模式(PAMP)”、“损伤相关分子模式(DAMP)”等,尽管这些术语普遍为人们所接受和使用,但其含义是否准确以及在多大程度上经得起时间考验仍是研究人员需重视的问题。
近日,来自法国波尔多大学的Thomas Pradeu和瑞士洛桑联邦理工学院(EPFL)的Bruno Lemaitre等在Immunity杂志上发表了一篇题为The conceptual foundations of innate immunity: Taking stock 30 years later的观点文章,他们在此重点回顾了固有免疫核心概念的发展并讨论了其适用范围。下面我们将对本文内容进行简要概述。

一、固有免疫如何识别病原体
1989年,Janeway首先提出PRR和PAMP的相关概念,他认为在基于基因重排的适应性免疫出现之前,免疫细胞即具有感知病原体一般分子结构模式(如脂多糖)的能力。模式识别受体(PRR)具有不同类型,分别是细胞质型、跨膜型和分泌型,目前已鉴定的PRR包括Toll样受体(TLR)、含有核苷酸结合结构域和富含亮氨酸重复序列的蛋白(NLRs)、RIG-I样受体和AIM2样受体等。然而,PRR检测系统存在一定的局限性,由于PAMP并非一成不变,肽聚糖的酰胺化或LPS的酰化等修饰能使病原体能逃避PRR的识别,部分微生物也缺乏经典的PAMP,此外,PRR-PAMP系统并不适用于系统发育上与其宿主接近的病原体,如难以检测后生动物体内的真核寄生虫。与这种依赖于微生物结构模式检测的PAMP触发免疫(PTI)不同,另一种效应触发免疫(ETI)基于对病原体编码的毒力因子或其活性的检测,从某种意义上来说,ETI更具有特异性。
一些免疫受体不能识别PAMP,但可以识别几乎所有宿主表达的分子模式,因此,缺乏这种“自我信号”的入侵者即能触发免疫反应。Karre于1986年提出了“丧失自我(missing-self)”假说,简而言之,宿主的所有有核细胞可组成型表达MHC-I,该分子与NK细胞表面受体结合抑制其功能,当细胞被病原体感染或发生癌变时,MHC-I表达下调进而活化NK细胞。另一个重要例子是NK细胞对在病毒感染时诱导的表达MICA和MICB等蛋白质的宿主细胞的检测,即对“改变自我(altered self)”的识别。
与感染无关的损伤相关分子模式(DAMP)也在固有免疫中发挥重要作用,例如存在于细胞内的肌动蛋白释放到胞外会触发JAK-STAT通路以协调损伤反应,DAMP也存在于植物中,一些病原体会分泌植物细胞壁降解酶,这些细胞壁片段以及受损细胞内的肽、核苷酸和氨基酸等会充当危险信号以激活防御反应,例系统素(systemin)。综上,PTI(PRR-PAMP系统)、ETI和DAMPs为不同的识别模式,PTI可理解为结构性识别,ETI是对病原体致病活动或效应因子的监视,而DAMP是响应非感染性损伤而释放的内源性宿主分子。
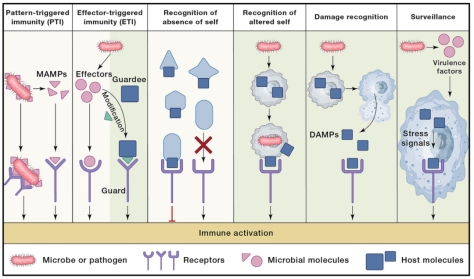 图1. 固有免疫的六种感知机制 图1. 固有免疫的六种感知机制
二、固有免疫效应机制
与适应性免疫及其“抗原特异性”相反,固有免疫长期以来被认为缺乏特异性,然而,许多固有免疫反应却是针对入侵病原体量身定制的。固有免疫携带有关病原体类别的信息及其他背景信息(如组织和亚细胞定位)来激活下游特定的信号转导途径,例如,果蝇中Toll和Imd途径分别被真菌和革兰氏阴性细菌选择性激活。相同的PAMP可诱发不同反应,具体取决于它们的定位,例如细胞外LPS、鞭毛蛋白和DNA都会与TLR结合并产生转录反应,而这些相同PAMPs的胞质检测则会导致炎症小体激活和细胞死亡。
尽管固有免疫效应器在动植物中具有多样性,但始终依赖于有限机制。第一类,专门针对病原体效应器,如阳离子抗菌肽可破坏比真核细胞摸带更多负电荷的细菌和真菌膜;第二类,不针对病原体,而是靶向其毒力因子并抑制其感染能力,如宿主为应对真菌蛋白酶而产生的蛋白酶抑制剂;第三类,PRR辅助清除机制,由可直接识别病原体的PRRs引导,补体蛋白水解产物C3b与病原体结合,再由表达C3b受体的吞噬细胞所吞噬;第四类可描述为“受感染细胞的自杀机制”,可由NK细胞诱导;第五类,可称为“营养免疫”,是隔离对病原体生长至关重要的代谢物,例如,在昆虫、脊椎动物和植物中,转铁蛋白或乳铁蛋白对铁的螯合限制了该元素的可用性,而该元素对于细菌生长是必不可少的。
 图2. 固有免疫效应机制。 图2. 固有免疫效应机制。
尽管长期以来人们一直认为专门的免疫细胞才具有免疫力,但最近越来越多的研究表明所有细胞都可以针对某些病原体(通常是病毒和细胞内细菌)发起“细胞内在”或“细胞自主”反应,例如抗菌肽或乳铁蛋白在各种上皮细胞中组成型表达。抗病毒限制因子在干细胞中组成型表达等,关于细胞内在免疫还有很多有待发现,这将构成未来研究的一个重要方向。
三、固有免疫记忆
免疫记忆是指宿主在一次感染后,对同一病原体的二次感染会变得更具有抵抗力,这是适应性免疫的核心特征,固有免疫是否具有记忆仍存在争议。该团队在文章中重新描述了固有和适应性免疫系统表现出的几种不同的记忆形式:第一类,感染原在宿主体内持续存在,原发感染导致低持续感染状态,提高了基础免疫力和继发感染生存率;第二类,感染原被清除,但初次感染产生的免疫效应物(如抗菌肽)持久存在;第三类,初次感染会触发持续的固有免疫反应,可能会提供针对继发感染的终生保护,与第二类不同,该类别中免疫反应不会减弱,例如细菌和古细菌中的CRISPR-Cas系统;第四类,初次免疫结束后恢复到基础水平再重新激活,引发更强的免疫反应,这一典例是脊椎动物中长寿命的记忆淋巴细胞。研究表明固有免疫系统也可以介导这种基于重新激活的记忆,例如信号分子表达增加可更快的诱导效应子,免疫细胞分化为“预激活状态”使继发感染反应更快。这种机制在植物中被称作“系统获得性抗性(SAR)”,即植物的局部组织被病原菌感染时发出可移动信号,在全身建立持久且广谱的抗病机制。Netea等人于2011年提出了另一个概念——“训练免疫”,他们在动物模型中观察到先前感染或接种细菌、真菌或蠕虫寄生虫可抵抗与适应性无关的异源感染,对后续感染的非特异性反应增加,这一概念的引入可能会进一步刺激对固有免疫记忆的研究。
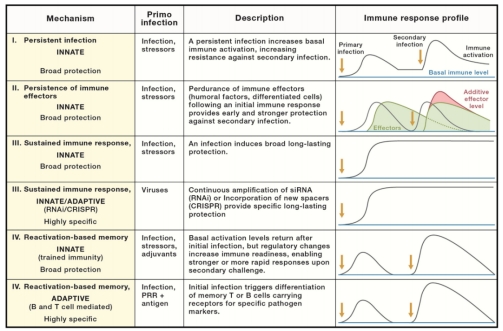 图3. 不同形式的免疫记忆 图3. 不同形式的免疫记忆
四、固有免疫耐受
免疫系统的一个关键特征是它们能够持续存在于宿主组织中为常驻微生物提供许可的生态位而不消灭它们。与长期以来认为自身免疫性疾病仅与适应性免疫有关的观点相反,一些自身免疫性疾病仅涉及固有免疫,例如在哺乳动物中,编码固有免疫受体(例如NLRC4或NLRP3)的基因突变会引起自身炎症综合征,对胞质核酸清除至关重要的基因(例如TREX1)突变可触发cGAS-STING固有免疫途径的组成型激活,导致Aicardi-Goutieres综合征或系统性红斑狼疮等疾病。
模式识别并不区分病原体和共生体,那么固有免疫系统如何处理肠道微生物组呢?最近发现了几种可能机制,首先,共生体被物理隔离在专门的生态位中,肠隐窝中抗菌提梯度建立了微生物“禁止入内”的区域,此外,PRR通常位于固有层中,只有当微生物突破这一屏障时才会发生反应。其次,宿主体内存在阻止免疫激活的负调节因子,内共生体释放的MAMP(微生物相关分子模式)引发的免疫反应会被负调节因子所抑制,豆科植物与根瘤菌的相互作用就是最好的例证。植物为细菌提供营养,而细菌通过将大气中的氮固定成植物可以使用的形式来获取氮,为了启动这种共生并抑制宿主免疫,细菌会分泌脂壳寡糖结瘤因子以被宿主细胞表面受体感知。
五、讨论
免疫现在被认为是所有生物中的普遍现象,传统上定义的基于淋巴细胞的适应性免疫仅是免疫研究领域中很小的一部分,大多数生物体都强烈依赖于固有免疫。
固有免疫以多种方式改变了免疫的核心概念,首先,免疫既是“系统性”,尤其会死“局部性”的,其次,尽管病原体是免疫系统进化中最核心的选择压力之一,,但免疫远远超出了对病原体的防御,固有免疫机制涉及组织稳态、共生和许多其他生理功能。截至目前,“特异性”、“适应性”、“记忆性”这些术语已不足以区分固有免疫和适应性免疫系统,因而需要在未来基于各种模式生物来重新概念化固有免疫。
原文链接:https://doi.org/10.1016/j.immuni.2024.03.007
|