植物DNA损伤识别和检查点控制的机理研究
植物DNA损伤反应(DDR)途径通过快速识别和修复DNA损伤来保护基因组完整性,如果不修复,可能会导致基因组不稳定。最常见的是,DNA修复伴随着短暂的细胞周期停滞,这使细胞能够在进行有丝分裂之前修复DNA损伤,但也因此影响植物的生长和产量。通过鉴定对DNA双链断裂或复制缺陷起反应的DDR蛋白和细胞周期调节因子,已经明确了这些蛋白和调节因子形成高度互联的网络。这些网络在转录和转录后水平上运行,包括液-液相分离和表观遗传机制。引人注目的是,尽管上游DDR传感器和信号元件在真核生物中非常保守,但一些更下游的效应元件在植物中却发生了分化,这可能是为了适应独特的生活方式特征。此外,DDR组分显示了古代植物物种、双子叶植物和单子叶植物的功能多样性。观察到的DDR突变体对铝毒性、磷酸盐限制和种子老化的抗性表明,了解植物DDR可能为应对气候变化的影响和相关的粮食安全风险提供解决方案.

随着世界人口持续增长和气候变化的影响变得更加明显,维护粮食安全是我们这个时代面临的最大挑战之一。气温上升、长期干旱和洪水对农作物产量和农业生产力产生了巨大影响。因为植物是不可移动的,它们暴露于这些新出现的极端和波动环境条件的一个后果是DNA损伤,这反过来影响细胞周期进程和细胞活力,从而影响生长和产量。事实上,许多外源性和内源性因素破坏了基因组的完整性,导致各种DNA损伤,如单链断裂(SSBs)和双链断裂(DSBs)、(氧化)碱基损伤、交联和嘧啶二聚体。幸运的是,DNA损伤会被一个高度复杂的信号网络迅速抵消,这个信号网络被称为“DNA损伤反应”(DDR)。完整功能和有效的DDR需要三个重要步骤:必须快速识别受损的DNA,启动信号级联(步骤1),该信号级联需要向效应器传输(步骤2)(步骤3)。这些效应物最终激活DNA修复,暂停细胞周期或通过杀死或推动细胞分化来阻止受影响细胞的增殖。尽管对生命树的广泛研究表明,许多DDR因子在真核生物中是保守的,但动物和植物之间的效应子和最终反应成分似乎有所不同。
在所有真核生物中,DDR途径的启动取决于DNA损伤的类型,位于两种磷酸肌醇3-激酶相关激酶的周围,即ATAXIA TELANGIECTASIA MUTATED (ATM)和ATAXIA TELANGIECTASIA AND RAD3-RELATED(ATR)。ATM主要由DNA DSBs激活,DNA DSB由MRE 11–RAD50–NBS1组成的蛋白质复合物识别。通过二聚化,该复合物将断裂末端拴在一起,并通过其内切和外切核酸酶活性切除DNA断裂侧丢失的5’末端,从而产生进一步修复所需的3’突出端。

图1.DDR的三步激活,实现高效的DNA修复。
NBS1亚基招募无活性的ATM二聚体,后者随后作为活化的ATM单体释放。
ATR主要由DNA SSBs和停滞的复制叉激活,也称为复制应激。在复制应激期间,由于没有新的链生物合成,DNA螺旋持续解链,单链DNA(ssDNA)片段出现。这种新出现的ssDNA由复制蛋白A和RAD17结合。复制蛋白A随后招募ATR相互作用蛋白,最终与ATR形成稳定的复合物。一旦ATR被激活,就会引发与DNA拓扑异构酶II结合蛋白的进一步结合,从而刺激ATR的自动激活,至少在酵母和人类细胞中是如此。RAD9–RAD1–HUS1复合物也可以激活ATR(图1)。
一旦被激活,哺乳动物ATM和ATR就会磷酸化肿瘤抑制蛋白p53,这是一种关键的DDR主调节因子,可触发DNA修复、细胞周期调节和凋亡的启动。尽管植物具有DNA损伤敏感成分的同源物(包括ATM和ATR),但它们缺乏p53,但它们依赖SUPPRESSOR OF GAMMA RADIATION 1(SOG1)。尽管这种植物特异性转录因子与p53没有任何序列相似性,但SOG1作为功能同源物,被ATM/ATR激活。磷酸化后,SOG1激活多种靶基因的表达,主要参与DNA修复和细胞周期检查点激活。除了SOG1,E2F转录因子和WEE1激酶是DDR的关键调节因子,构建了在转录和转录后水平上运行的复杂调控网络。总之,这些网络提供了一种质量控制机制,防止DNA受损的细胞进入有丝分裂。
在该综述中,作者向我们总结了DNA损伤感知和修复机制:包括RAD51的募集和功能,5/6号染色体的结构维护;DNA损伤检查点控制机制;作物和古代物种中的DDR,以及DDR在应对环境压力方面的重要性。

图2.RAD51核丝的多步骤形成和活性SMC5/6复合物。
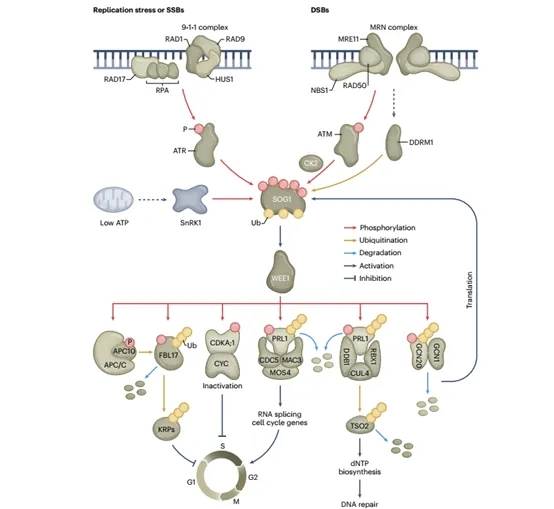
图3.SOG1/ WEE1介导的DNA损伤检查点的分子机制
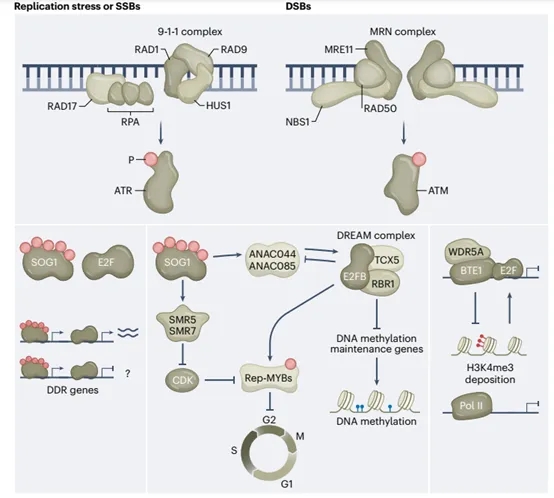
图4.DREAM复合物在DNA损伤检查点控制中的作用
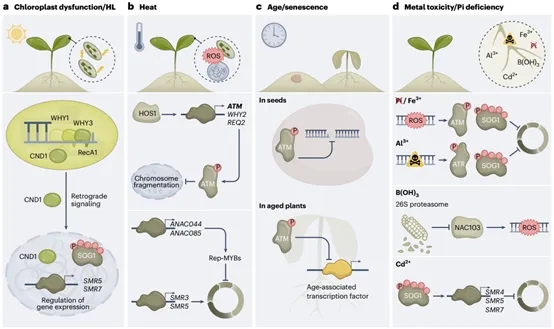
图5.DDR功能在各种极端环境中的重要性
|