|
无论在原核生物中还是在真核生物中,新合成的蛋白质必须转运到特定的亚细胞位置或运输到胞外才能发挥其相应活性,这种现象称为
蛋白质导向。
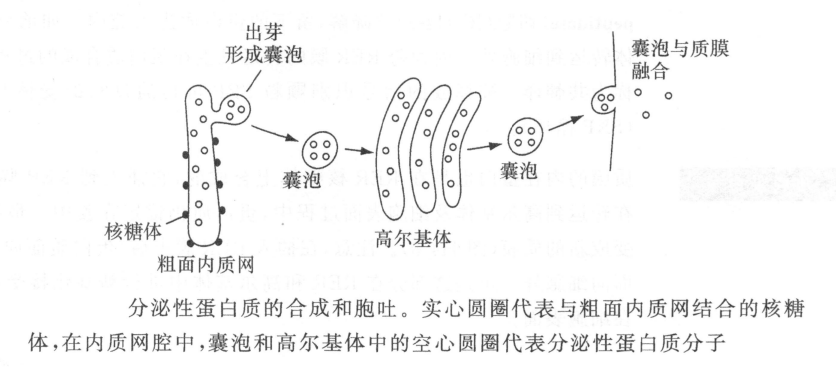
分泌性蛋白质有一段N端信号肽,它引导蛋白质在粗面内质网(RER)上合成。合成时,蛋白质穿过粗面内质网膜进入腔内。然后由RER出芽形成囊泡,将合成的蛋白质运输到高尔基复合体进行蛋白质糖基化。接着由其他的囊泡将蛋白质运到细胞质膜上,运输囊泡与质膜融合将蛋白质释放到细胞外。
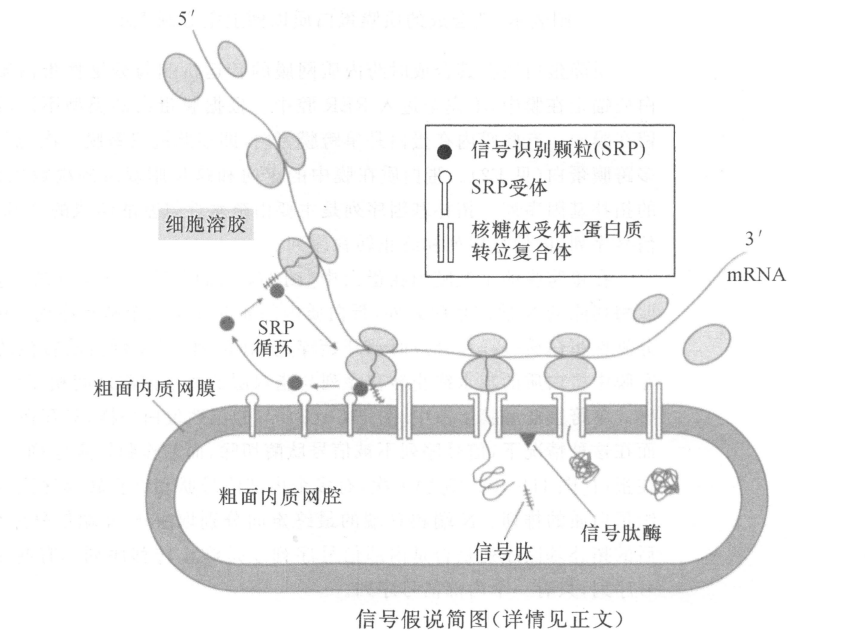
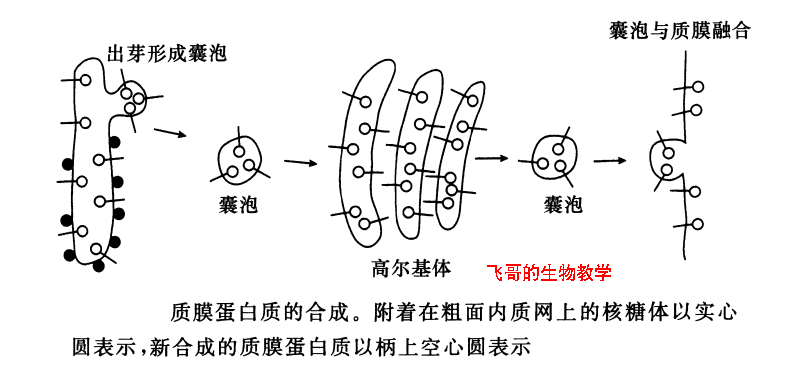
质膜蛋白质也在RER上合成并插入RER膜(最终成为细胞质膜),不是释放到RER 腔中。质膜蛋白质有的只有一次跨过质膜(I型和Ⅱ型膜内在蛋白),有的可多次跨膜(Ⅲ型膜内在蛋白)。蛋白质在膜中的取向由多肽链中的拓扑遗传序列所决定。I型膜蛋白有可被切割的 N 端信号序列和一段疏水性的终止转移序列,Ⅱ型有一段不可被切割的信号序列兼作膜锚定序列,Ⅲ型有多个信号序列和终止转移序列。通过糖基磷脂酰肌醇(GPI)锚定在膜上的蛋白质具有被切割的N端信号序列和连接GPI锚钩的C端疏水性的序列。
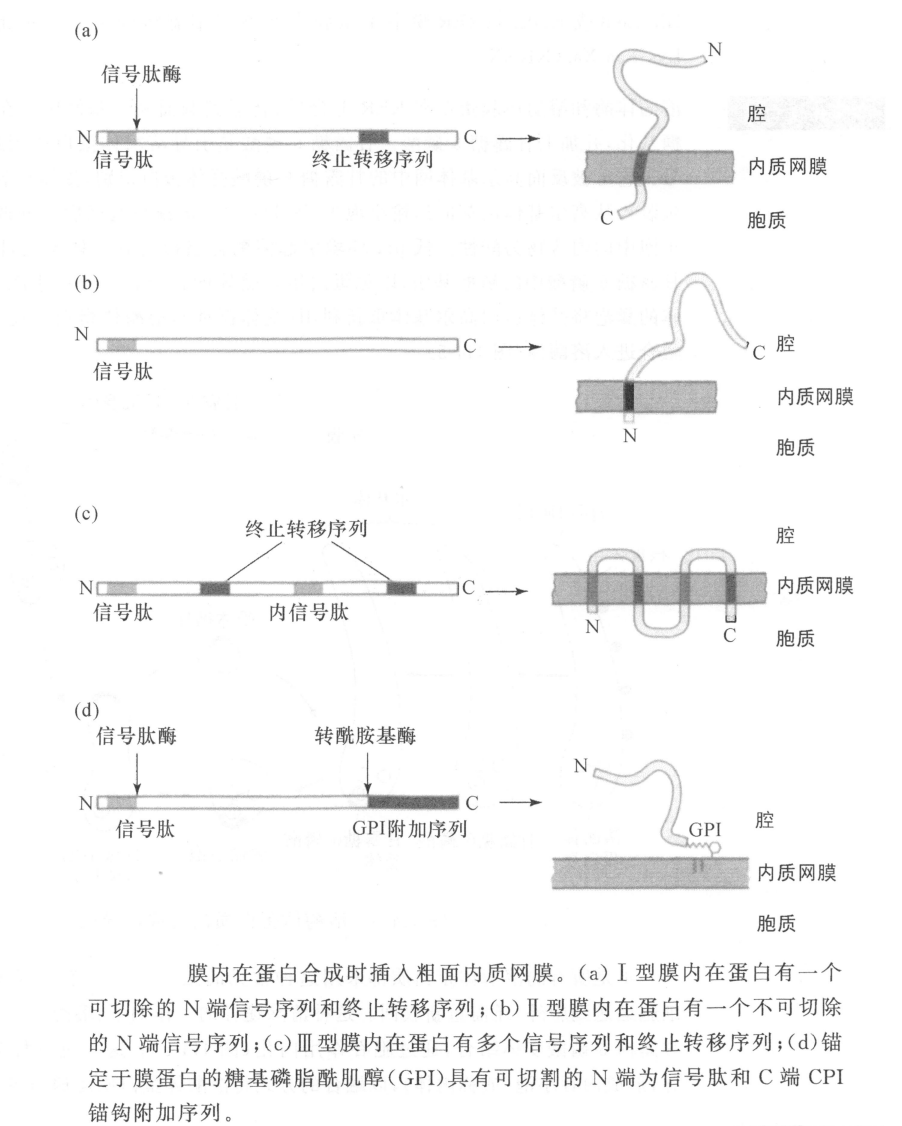
定位在内质网的蛋白质有N 端信号序列,在粗面内质网上合成,进入腔内或插入粗面内质网膜中,C端氨基酸序列(可溶性内质网腔蛋白为KDEL,I型膜内在蛋白为KKXX)被特异性受体蛋白识别,将蛋白质保留在内质网中。
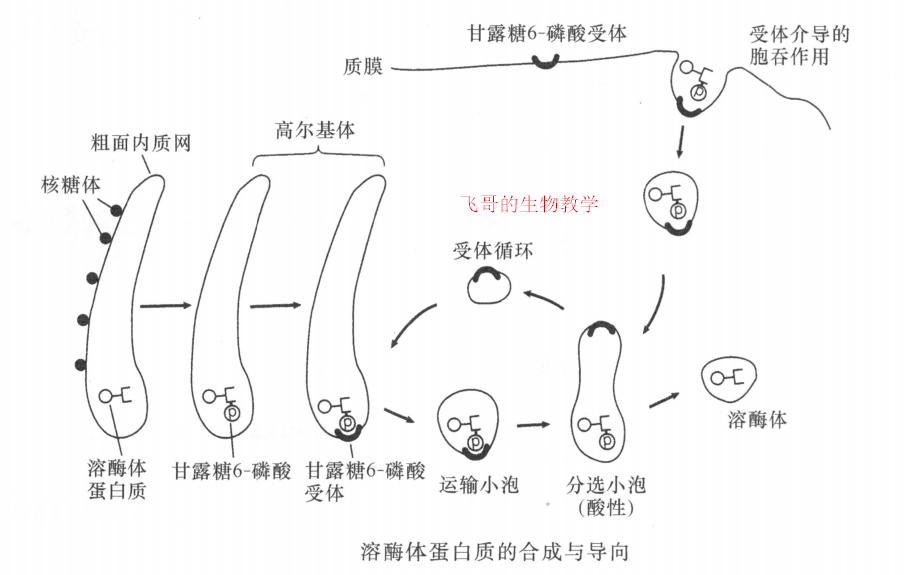
定位于溶酶体的蛋白质在顺面高尔基网中加入甘露糖6-磷酸信号,并被反面高尔基网中的受体蛋白识别。溶酶体蛋白质即被特异的囊泡运输到晚期内体中,成熟后进入溶酶体。6-磷酸甘露糖受体返回高尔基体中重复使用。
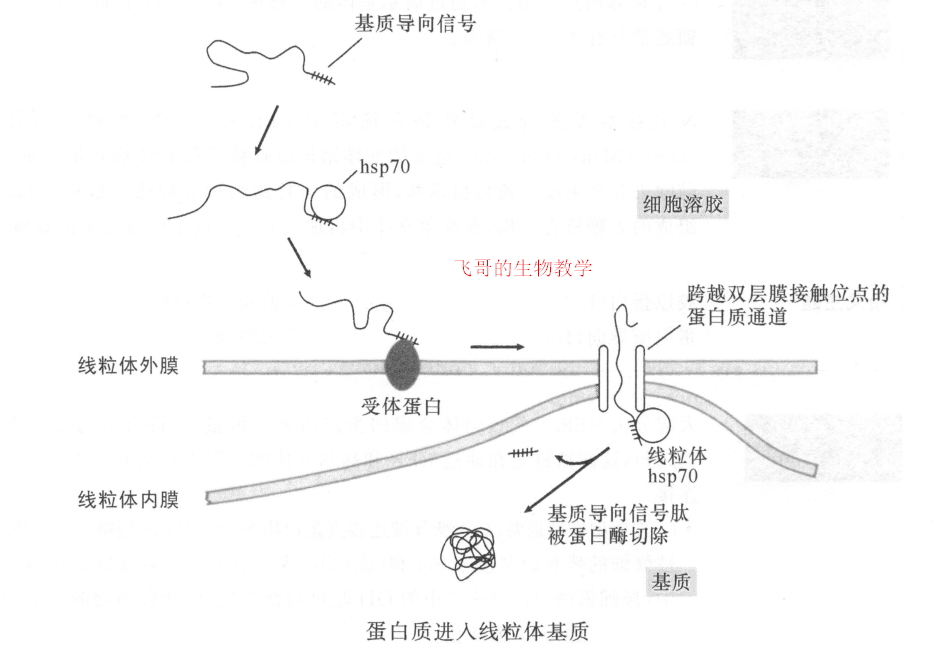
大部分线粒体和叶绿体蛋白质在细胞溶胶游离的核糖体上合成后,释放到细胞溶胶中后再进入细胞器。进入到线粒体基质的蛋白质需要基质导向序列,摄入的部位在线粒体内膜和外膜的接触处。该过程由 hsp70 和 hsp60介导,并需要水解 ATP 和线粒体内膜两侧的电化学梯度。定位于线粒体其他部位或叶绿体的蛋白质需要两种信号。
定位于细胞核的蛋白质需要一个典型的核定位信号,信号长度为4~8个氨基酸,位于蛋白质内部。蛋白质经核孔进入细胞核中,需要ATP水解。
|
|