1 内质网的形态结构与类型
内质网(endoplasmic reticulum, ER)是一个连续的膜性三维管网结构系统,广泛分布于除成熟红细胞以外的所有真核细胞的胞质中。它可与高尔基体、溶酶体等内膜系统的其他组分转换,还可与细胞核外膜直接连通。
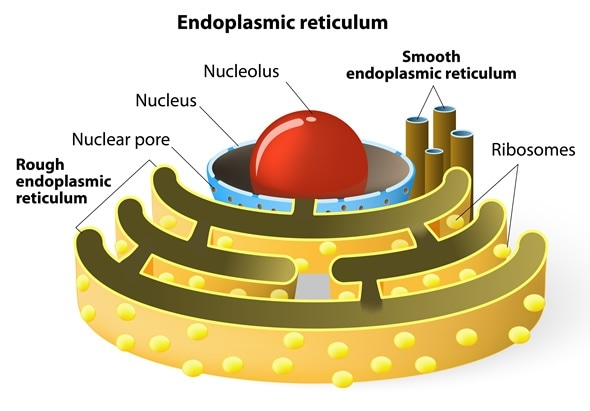
内质网可划分为两种基本类型:
-
糙面内质网(rough endoplasmic reticulum, RER)
-
-
-
主要与外输性蛋白质和多种膜蛋白的合成、加工、转运有关
-
光面内质网(smooth endoplasmic reticulum, SER)
2 内质网的化学组成
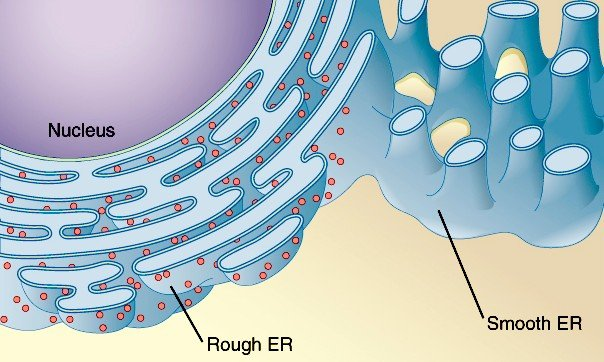
内质网以脂类和蛋白质为其结构的主要组成成分。膜脂含量约30%~40%,蛋白质含量在60%~70%之间。
内质网含有多种酶蛋白,大致可划分为以下几种类型:
-
-
-
与碳水化合物代谢功能相关的酶类,如葡萄糖-6-磷酸酶。
-
3 内质网的功能
3.1 糙面内质网
糙面内质网的主要功能是进行蛋白质的合成、加工修饰、分选及转运。
3.1.1 信号肽指导分泌蛋白在糙面内质网合成
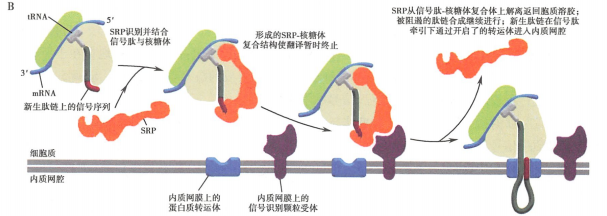
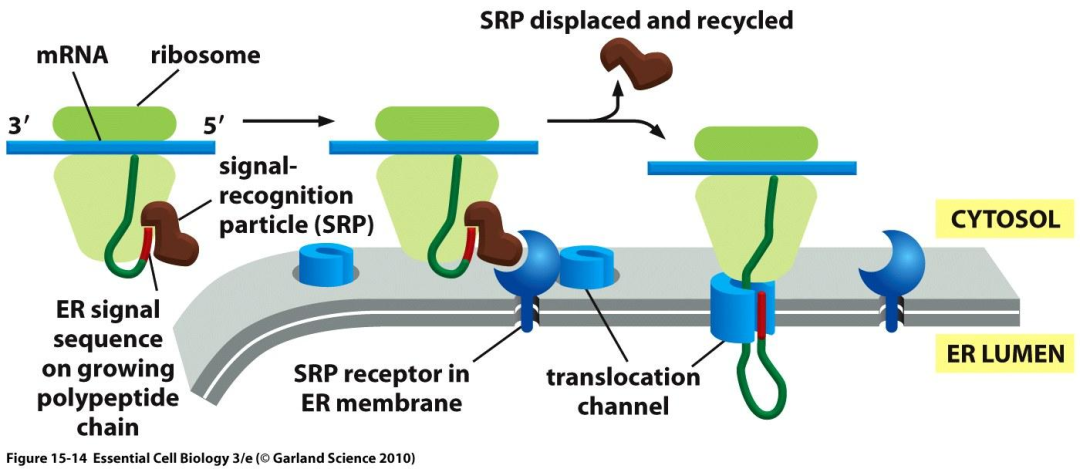
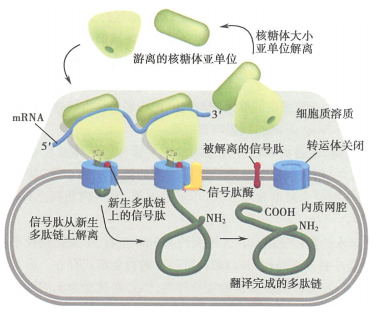
所有蛋白质的合成都起始于细胞质基质中游离的核糖体上。分泌蛋白(secreted protein)多肽链在其合成起始后不久随核糖体一起附着于糙面内质网上,不断延伸的多肽链穿过内质网膜直至肽链合成完成。
信号肽(signal peptide)是普遍存在于分泌蛋白肽链N端的一段特殊的疏水氨基酸序列,是指导多肽链在糙面内质网上进行合成的决定因素。
核糖体结合内质网、肽链穿越内质网膜,还有赖于细胞质基质中的信号识别颗粒(signal recognition particle, SRP)、内质网膜上的信号识别颗粒受体(SRP-receptor, SRP-R,又称停靠蛋白(docking protein))、被称之为转运体(translocon, translocator)的易位蛋白质。
-
新生的分泌蛋白在细胞质基质中的游离核糖体上起始合成
-
新生肽链N端的信号肽被翻译后可立即被细胞质基质中的SRP识别、结合,SRP另一端结合于核糖体上,翻译暂时终止
-
与信号肽结合的SRP识别、结合内质网膜上的SRP-R,并介导核糖体附着于内质网膜的转运体上
-
-
在信号肽的介导下,合成中的肽链通过由核糖体大亚基的中央管和转运体共同形成的通道穿膜进入内质网网腔
-
信号肽序列被内质网膜腔面的信号肽酶切除,新生肽链继续延伸
-
完成肽链合成的核糖体大、小亚基解聚,并从内质网上脱离
-
3.1.2 新生多肽链的折叠与装配
-
内质网腔中丰富的氧化型谷胱甘肽(GSSG)和蛋白二硫键异构酶有利于二硫键形成
-
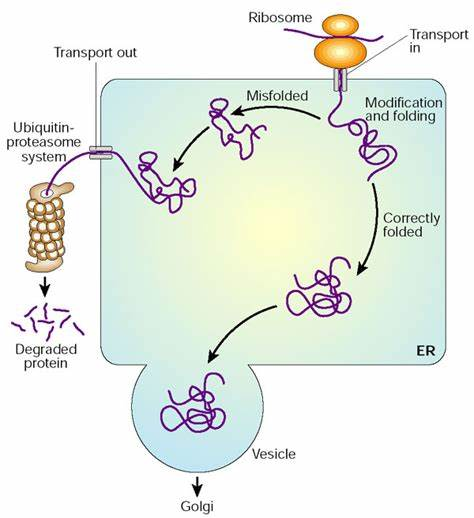
分子伴侣(molecular chaperone)通过与多肽链的识别结合来协助它们的折叠组装和转运,包括使折叠错误的多肽、尚未完成装配的蛋白亚单位驻留
内质网腔内未折叠蛋白的积聚,可通过未折叠蛋白反应(unfolded protein response, UPR)使内质网分子伴侣表达升高,从而有利于蛋白质的正确折叠和组装。
3.1.3 蛋白质的糖基化
发生在糙面内质网的糖基化主要是寡糖与蛋白质Asn残基侧链上氨基基团的结合(N-连接)。
3.1.4 蛋白质的胞内运输
由附着核糖体合成的各种外输性蛋白质,经过在糙面内质网中的修饰、加工后,最终被内质网膜包裹,以“出芽”方式形成膜性小泡而转运。
-
途径一:进入高尔基体进一步加工浓缩,最终以分泌颗粒的形式被排吐到细胞外
-
途径二:(仅见于某些哺乳动物的胰腺外分泌细胞)以膜泡形式直接进入大浓缩泡,进而发育成酶原颗粒,然后被排出细胞
3.2 光面内质网
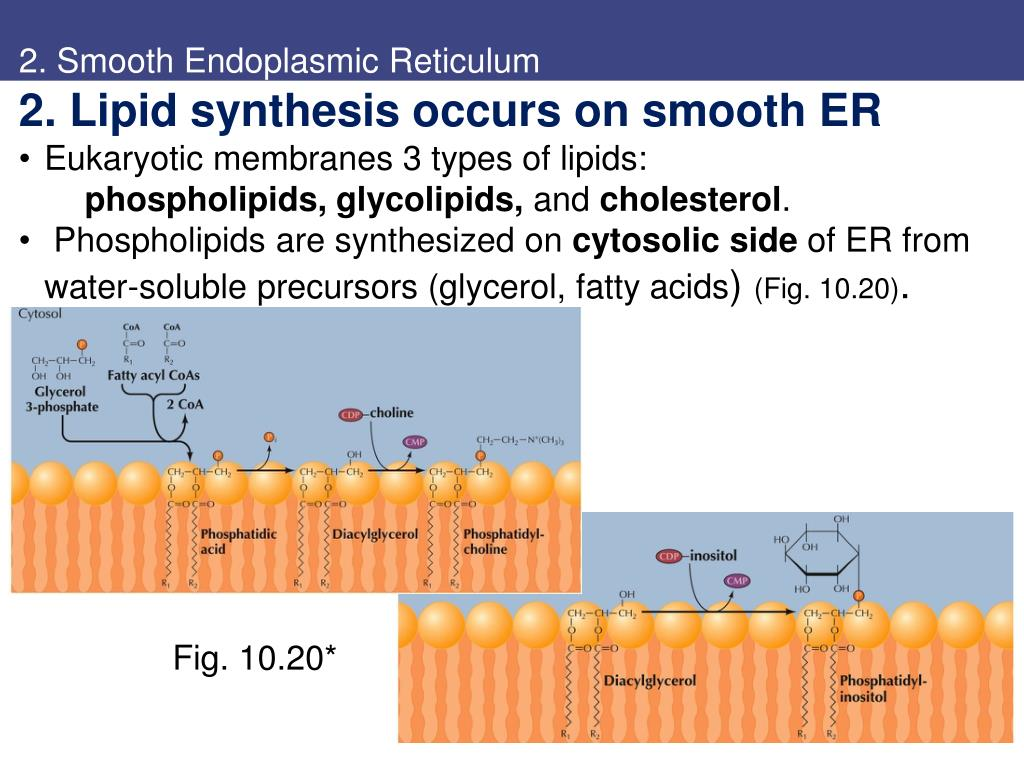
3.2.1 光面内质网参与脂质的合成和转运
经由小肠吸收的脂肪分解物甘油、甘油一酯和脂肪酸进入细胞后,在内质网中可被重新合成为甘油三酯。在光面内质网合成的脂类常常会与来自糙面内质网的蛋白质结合形成脂蛋白,然后经由高尔基体分泌。
在类固醇激素分泌旺盛的细胞,其发达的光面内质网中存在着与类固醇代谢密切相关的关键酶。
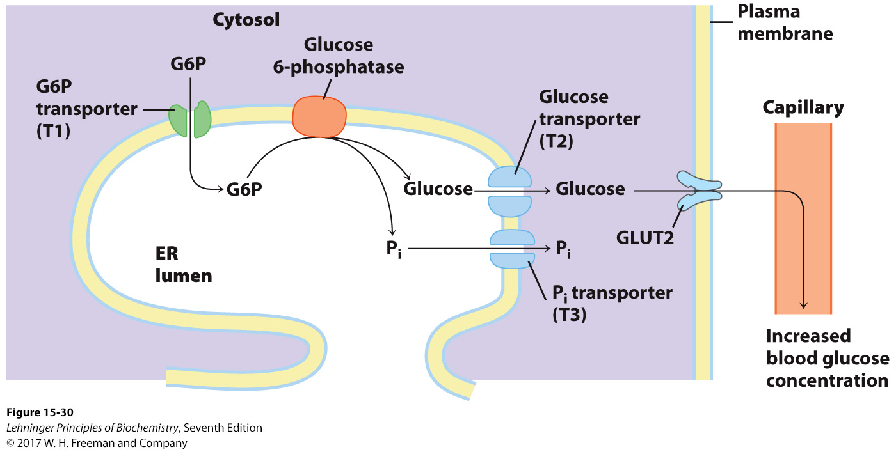
3.2.2 光面内质网参与糖原的代谢
内质网参与糖原的分解过程。肝细胞光面内质网网腔面存在葡萄糖-6-磷酸酶等,胞质面存在糖原磷酸化酶等。
3.2.3 光面内质网是细胞解毒的主要场所
肝脏的解毒作用主要由肝细胞中的光面内质网来完成,含有丰富的氧化及电子传递酶系(成为加单氧酶系(monooxygenase))。

解毒的基本机制是:在电子传递的氧化还原过程中,通过催化多种化合物的氧化或羟化,一方面使得毒物、药物的毒性作用被破坏,另一方面增强化合物的极性使之易于排泄。
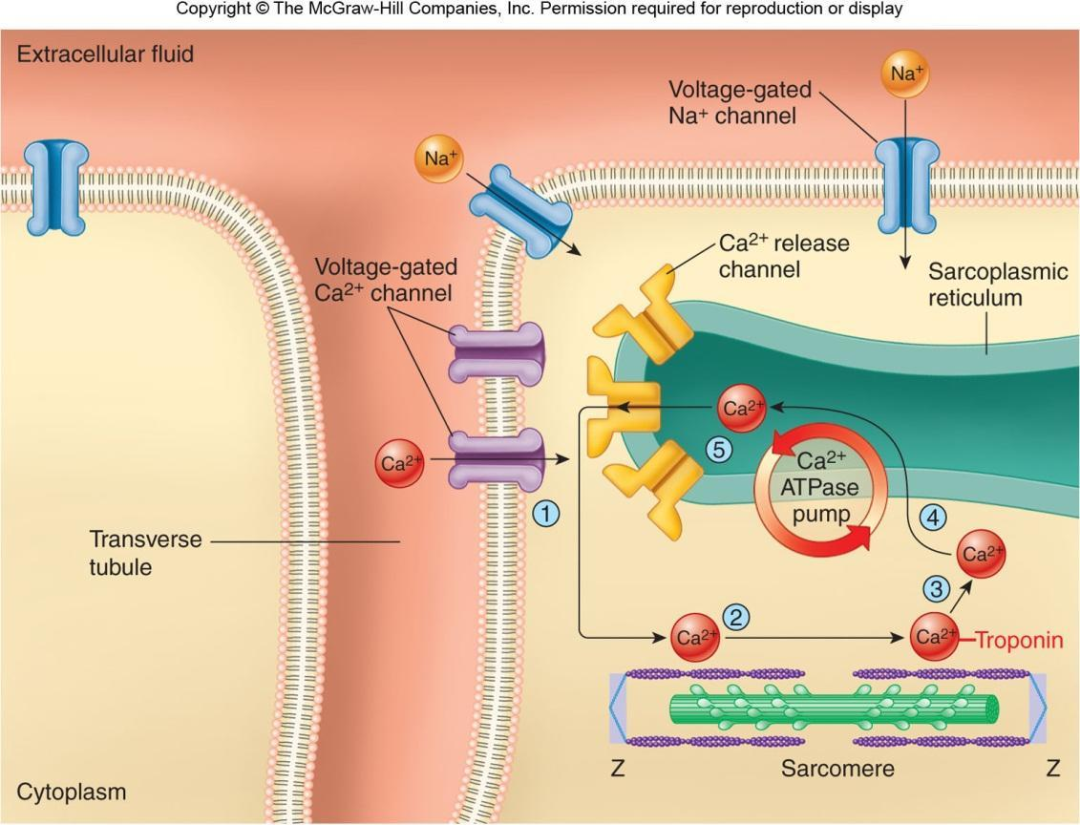
3.2.4 光面内质网是肌细胞Ca2+的储存场所
肌细胞中十分发达的光面内质网特化为肌质网(sarcoplasmic reticulum)。肌质网膜上的Ca2+-ATP酶把细胞质基质中的Ca2+泵入网腔储存,受到刺激时引起Ca2+向细胞质基质释放。
|