2.3 高尔基体的结构和功能
高尔基体排列较为整齐,由4-8层扁平潴泡平行堆叠成的扁平膜囊堆和周围小泡构成。它在细胞中往往有比较恒定的位置和方向,是一种有极性的细胞器,靠近细胞核的一面膜囊弯曲成凸面,称为形成面或顺面,面向质膜一面常成凹面,又称成熟面或反面。高尔基体可分为几个不同的功能区室(表2)。
高尔基体的主要功能是参与蛋白质的分选与分选蛋白(包括膜脂)的转运,伴随此过程,发生N-连接寡糖链的加工、部分蛋白质的O-连接糖基化修饰,以及鞘磷脂与糖脂的合成等。

3 分泌蛋白的合成与运输
3.1分泌蛋白的分选转运途径
真核细胞分泌蛋白分选途径为共翻译转运途径,即蛋白质合成起始后,在信号肽的指引下一边翻译,一边进入粗面内质网腔中,随后经高尔基体加工、分选后分泌到细胞外(图2)。分泌蛋白的分选转运基于膜泡转运,即蛋白质包裹在不同类型的囊泡中,从粗面内质网合成部位定向转运至CNG,再从TNG转运至质膜并分泌到细胞外,该过程涉及供体膜与靶膜的识别与融合。
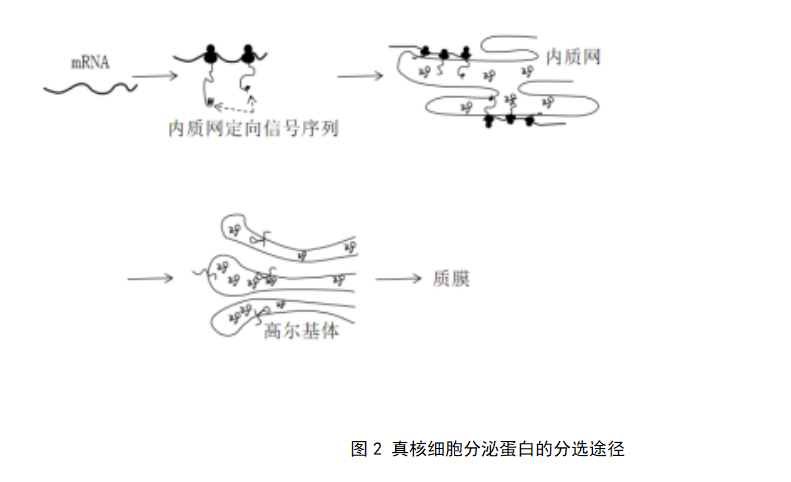
3.2 信号假说与分泌蛋白的合成
3.2.1 信号假说
信号假说认为,分泌蛋白的合成始于细胞质中的游离核糖体,新生肽N端有一段16--26个氨基酸残基组成的具有信号作用的序列,称为信号序列或称为信号肽,负责指导蛋白质一边合成一边通过转运蛋白水性通道进入内质网腔,信号肽完成任务后被切除,完全合成的新生肽进入内质网腔,转运蛋白关闭,直到有新的核糖体结合才打开。
3.2.2 分泌蛋白的合成
信号假说得到许多实验的证实,分泌蛋白合成步骤如下:①细胞质基质中游离核糖体大小亚基结合后起始合成分泌蛋白。②肽链延伸。③肽链延伸至80个氨基酸左右时,N端的内质网信号序列暴露出核糖体。④信号肽与信号识别颗粒(signal recognition particle,SRP)中的信号序列结合位点识别与结合;同时,SRP中的翻译暂停结构域暂时中止肽链合成。⑤SRP与内质网膜上的SRP受体结合,SRP-信号序列-核糖体-mRNA复合物锚定到内质网膜,SRP受体水解GTP的同时释放SRP,并打开内质网膜上的蛋白质转运通道,核糖体与通道结合,新生肽N端信号序列插入转运蛋白的水性通道,肽链又开始延伸。⑥释放的SRP将返回细胞质基质中去介导另一个新生多肽及核糖体与内质网膜的结合。⑦在信号肽酶的作用下切除信号肽。⑧分泌蛋白被释放到内质网的腔中。⑨核糖体大小亚基彼此分离。分泌蛋白合成过程中肽链输送至内质网的流程见图3。
3.3 分泌蛋白的修饰加工
3.3.1 分泌蛋白在内质网的折叠与修饰
当肽链在信号序列的指引下进入内质网腔后,这一信号序列任务完成,随即会被信号肽酶切除,分泌蛋白被释放到内质网腔中。随后肽链在内质网腔内进行形式多样的修饰加工,包括膜蛋白的分选、蛋白质的折叠装配、蛋白质的糖基化修饰、羟基化修饰、脂酰基化修饰及二硫键的形成以及蛋白质的折叠质量的监控等。许多分泌蛋白含有二硫键,内质网中的二硫键异构酶催化新生肽链二硫键的形成,以及通过异构化消除不正确的二硫键,它能加速蛋白质折叠。内质网中的分子伴侣在新生肽进入内质网腔的转移过程中起作用,也可帮助多肽正确折叠,防止新生肽链在转运过程中断裂或变性以及多肽错误折叠和聚合。内质网中进行的N-连接的糖基化修饰也有利于蛋白质的正确折叠。错误折叠的多肽链在分子伴侣的帮助下从它们进入内质网腔的路线返回细胞质基质。
3.3.2 分泌蛋白在高尔基体中的修饰加工
CNG接受来自内质网的蛋白质并将其分类后转入高尔基体中间膜囊进行修饰加工。不同蛋白质的酶解加工方式不同,可归纳为以下几种类型:①无生物活性的蛋白原可在高尔基体中将其两端序列或N端切除,形成成熟多肽,如胰岛素、胰高血糖素。②有些蛋白质分子在粗面内质网中合成时便是含有多个相同氨基酸序列的前体,经过在高尔基体中水解,形成同种有活性的多肽,如神经肽。③一个蛋白质分子的前体中含有不同的信号序列,最后加工成不同的产物。
3.3.3 分泌蛋白修饰加工的实例
分泌蛋白的修饰加工,可结合胰岛素的形成进行理解。在核糖体上合成由110个氨基酸组成的前胰岛素原,在N端有一段信号序列,即信号肽。信号肽引导新生多肽进入内质网腔后,立即被酶切除,剩余的多肽链折叠成含3个二硫键的胰岛素原。后者进入高尔基体,在酶的催化下除去一段连接胰岛素A链和B链的连接肽(C肽),转变为胰岛素(图4)。
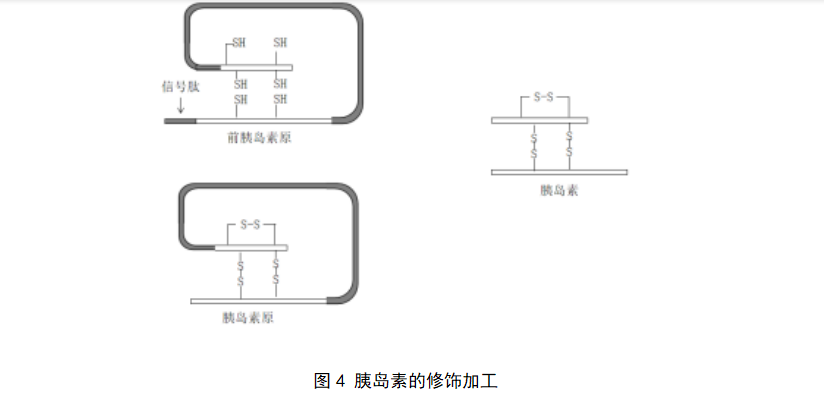
3.4 分泌蛋白质的运输
分泌蛋白主要通过运输小泡在细胞器及细胞表面转运,其运输可归纳为三个阶段:供体膜出芽产生小泡、小泡转运、小泡到达靶膜并融合,卸载货物。已知有三种类型的有被小泡:COP I有被小泡、COPⅡ有被小泡、网格蛋白有被小泡。COP I有被小泡可介导CNG到内质网的运输、TNG到质膜的非选择性运输以及高尔基体中间膜囊间的运输。COPⅡ有被小泡主要介导内质网出芽输出的蛋白质到CNG。网格蛋白有被小泡承担选择性的物质运输,从TNG出芽形成的分泌小泡,介导溶酶体酶的运输,也可介导由质膜内吞形成的选择性内吞泡。
本文重点介绍COPⅡ有被小泡介导的从内质网输出的蛋白质到CNG的分泌,TNG到质膜的运输不再赘述。内质网加工后的分泌蛋白需装载到COPⅡ小泡,这些小泡在内质网出口位点出芽形成。COPⅡ有被小泡有两层包被,内层由Sec23/Sec24组成,外被由Sec13/Sec31组成。COPⅡ有被小泡的装配需要的包被招募GTPase是Sar1,Sar1通常以结合有GDP的非活性状态存在于细胞质。当内质网膜开始形成COPⅡ被膜时,内质网膜上的特异性蛋白Sar1-GEF的细胞质结构域与Sar1结合,释放GDP的同时结合GTP,导致构象改变,暴露出两亲性螺旋进而插入到内质网膜的细胞质侧的膜小叶(图5-a)。之后Sar1-GTP将COPⅡ的衔接蛋白Sec23、Sec24招募到膜上,构成包被的内层,Sec24可与内质网膜上

