| |
| 【教学参考】8种细胞器的结构和功能 |
|
| 来源:仪征中学 时间:2023-10-20 |
| |
|
1.核糖体的结构和功能
核糖体是一种颗粒状的结构,没有被膜包裹,其直径为
25 nm
,主要成分是蛋白质与
rRNA
。蛋白质含量约占
40%
,
rRNA
约占
60%
。核糖体蛋白分子主要分布在核糖体的表面,而
rRNA
则位于内部,二者靠非共价键结合在一起。
在真核细胞中很多核糖体附着在内质网的膜表面,成为附着核糖体。
在原核细胞的细胞膜内侧也常有附着核糖体。
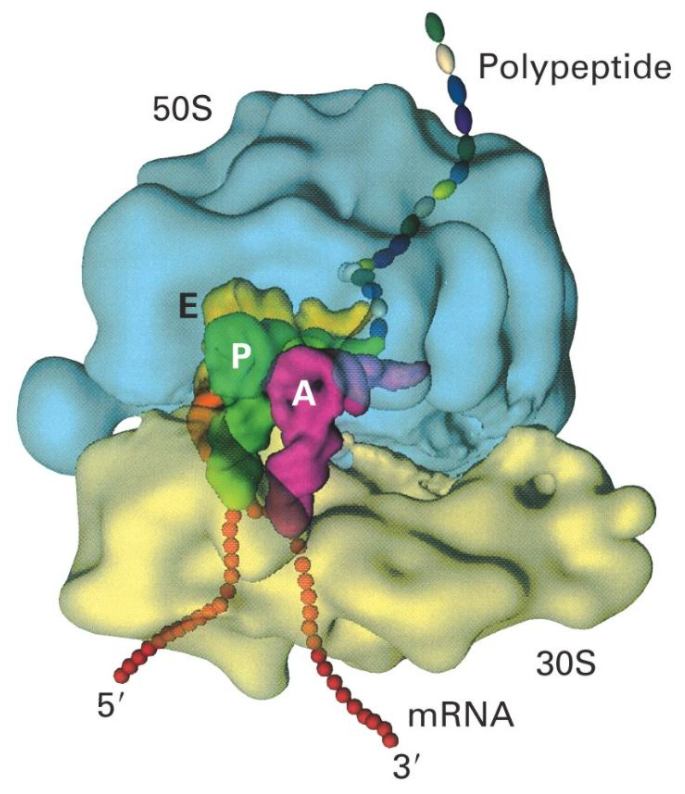
还有些核糖体不附着在膜上,而呈游离状态,分布在细胞质基质内,称为游离核糖体。附着核糖体和游离核糖体所合成的蛋白质种类不同,但核糖体的结构和化学组成是完全相同的。
细胞内有两种类型的核糖体:一种是
70S
的核糖体,原核细胞中的核糖体属于此类,真核细胞线粒体与叶绿体内的核糖体也近似于
70S
;另一种是
80S
的核糖体,存在于真核细胞中。不论
70S
或
80S
核糖体,均由大小不同的两个亚单位构成。
70S
核糖体可解离为
50S
和
30S
两个大小亚单位,
80S
核糖体可解离为
60S
和
40S
两个大小亚单位。
核糖体是合成蛋白质的细胞器,其功能是按照
mRNA
的指令把氨基酸高效且精确地合成肽链。核糖体在细胞内并不是单个独立地执行功能,而是由多个甚至几十个核糖体串连在一条
mRNA
分子上,高效地进行肽链合成。这种具有特殊功能与形态结构的核糖体与
mRNA
聚合体称为多聚核糖体。在真核细胞中每个核糖体每秒能将两个氨基酸残基加到肽链上;而在细菌细胞中,每秒可将
20
个氨基酸残基加到肽链上,因此合成一条完整的肽链平均需要
20s
到几分。
内质网是由
Porter
等人在
1945
年发现的。他们利用电镜在成纤维细胞中观察到一些形态和大小略有不同的网状结构,并集中在内质中,因此将这些结构称为内质网。
内质网是由一层膜形成的囊状、泡状和管状结构,并形成一个连续的网膜系统。
内质网通常占细胞的生物膜系统的一半左右,占细胞体积的10%以上
。根据内质网上是否附有核糖体,将内质网分为两类,即粗面内质网和光面内质网。粗面内质网多呈大的扁平膜泡,排列整齐。它是核糖体和内质网共同组成的复合结构,普遍存在于细胞中,特别是合成分泌蛋白的细胞。在结构上,粗面内质网与细胞核的外层膜相连。无核糖体附着的内质网称为光面内质网,通常为小的管状和小的泡状,广泛存在于各种类型的细胞中。光面内质网是脂质合成的重要场所。
内质网可通过出芽方式,将合成的蛋白质或脂质转运到高尔基体。
高尔基体是意大利科学家高尔基(
C.Golgi
)在
1898
年发现的。它是普遍存在于真核细胞中的一种细胞器。在电镜下观察到,高尔基体是由一些排列较为整齐的扁平膜囊堆叠在一起,构成了高尔基体的立体结构。扁平膜囊多呈弓形,也有的呈半球形,均由光滑的膜围绕而成。在扁平膜囊外还包括一些小的膜泡。整个高尔基体结构分为形成面和成熟面,来自内质网的蛋白质和脂质从形成面逐渐向成熟面转运。
高尔基体与细胞的分泌功能有关,能够收集和排出内质网所合成的物质,它也是聚集某些酶原的场所,参与糖蛋白和黏多糖的合成。高尔基体还与溶酶体的形成有关,并参与细胞的胞吞和胞吐作用。
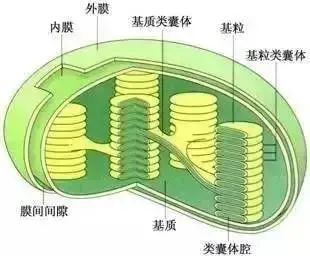
叶绿体是植物进行光合作用的细胞器。具有叶绿体的植物除高等植物外,还有真核藻类。叶绿体的形状因物种的不同而有所差异。藻类的叶绿体形态差异较大,可以是板状、带状、杯状、囊状、星状等。高等植物的叶绿体一般形状比较固定,多为扁平的椭球形,平均直径为
4~6 μm
,厚
2~3μm
。
叶绿体由双层膜包被,每层膜厚
6~8 nm
,外膜与内膜之间有
10~20 nm
宽的膜间隙。两层膜均由单位膜(由脂质双层及嵌合蛋白质构成的一层生物膜)组成,具有选择透过性。叶绿体膜内的基础物质称为基质。基质中悬浮着复杂的膜片层系统,其基本单位是由单位膜封闭形成的扁平小囊,称为类囊体(也称片层)。类囊体有规律地垛叠在
一
起形成好似一摞硬币的结构被称为
基粒类囊体。贯穿在两个或两个以上基粒之间没有发生垛叠的类囊体,称为基质类囊体。
相邻基粒由基质类囊体链接在一起,使类囊体腔之间彼此相通,因而,一个叶绿体内的全部类囊体实际上是一个连续的封闭的三维结构。类囊体膜上有多种蛋白复合体,包括光合电子传递体和光合色素蛋白质复合体,是光合作用中进行光反应的结构。类囊体膜上的光合色素负责在光合作用中吸收光能,这种膜片层系统极大地增加了光合作用中的受光面积,提升了光合作用的效率、叶绿体的基质中有可溶性的蛋白(酶)以及其他活跃的代谢物质,其中包括光合作用中催化碳固定的酶系统,因此,光合作用中二氧化碳的固定、还原是在叶绿体基质中完成的。基质中还存在叶绿体自身的
DNA
、
RNA
和核糖体,能够自主进行遗传物质的传递以及蛋白质的合成。
与叶绿体相比,线粒体要小一些,直径
0.5~1μm
,长
1~2 μm
,通常呈椭球状或圆柱体。线粒体也由内外两层单位膜包裏,内外膜之间有腔。外膜平整光滑,内膜内折形成嵴。内膜上分布有许多规律排列的带柄的球状小体,即
ATP
合酶,它利用电子传递过程中形成的质子跨膜电化学势梯度合成
ATP
。线粒体膜的内部为基质,具有一定的
pH
和渗透压,含有进行三羧酸循环等多种代谢途径的酶。除此之外,线粒体基质中还含有自身的
DNA
、
RNA
和核糖体。
叶绿体和线粒体有自己的基因组、蛋白质合成系统和膜系统,说明这两种细胞器具有自我繁殖所必须的基本物质,能够进行转录和翻译,这就保证了它们在真核细胞中仍然有一定程度的自主性。有关线粒体遗传的研究,已经清楚地显示出了线粒体的自主性。
通过细胞的结合,一种细胞的线粒体可以在杂种细胞及其后代的体内生活和增殖。不仅一种动物的线粒体可以生活在另一种动物的细胞中,而且连叶绿体都可以人工地使它们生活在动物细胞中。例如,在离体的小鼠成纤维细胞的培养基中加入菠菜叶绿体,半小时就可以看到有些叶绿体已经在小鼠的细胞中生存。事实上,叶绿体在活体外也能独自生存相当长的时间。例如,把刺海松(一种管藻)的叶绿体培养在简单的无机培养基中,5d后它们仍然能够进行光合作用。
迄今为止,已知线粒体基因组仅能编码约
20
种线粒体膜和基质蛋白质,并在线粒体自身的核糖体上合成,叶绿体能够自身编码合成的蛋白质比线粒体多一些,但也仅有
60
多种。然而,参与组成线粒体和叶绿体的蛋白质各有上千种之多,其中绝大多数的蛋白质仍然是由核基因编码,在细胞质核糖体上合成后转移到线粒体与叶绿体当中的。细胞核与发育成熟的线粒体或叶绿体之间存在着密切的、准确的严格调控的协同机制。也就是说,线粒体和叶绿体的自主程度是有限的,它们对核遗传系统具有很强的依赖性。所以,线粒体和叶绿体被称为半自主性细胞器。
溶酶体是动物细胞中一种由膜构成的细胞器,呈小球状,外面由一层非渗透性单位膜包被。溶酶体是一种动态结构,它不仅在不同类型细胞中形态大小不同,而且在同一类细胞的不同发育阶段形态大小也不相同。溶酶体的主要功能是
消化作用
,其消化底物的来源有三种途径:一是自体吞噬,吞噬的是细胞内原有物质;二是吞噬体吞噬的有害物质;三是胞吞吞人的营养物质。溶酶体除了具有吞噬消化作用外,还具有
自溶作用
,即某些即将衰老的细胞靠溶酶体破裂释放出各种水解酶将自身消化。此外,溶酶体内的酶也可释放到细胞外,对细胞外基质进行消化。根据溶酶体处于完成其生理功能的不同阶段,大致可分为初级溶酶体和次级溶酶体。初级溶酶体是刚刚从高尔基体形成的小囊泡,仅含有水解酶类,但无作用底物,而且酶处于非活性状态。次级溶酶体中含有水解酶和相应的底物,是一种将要或正在进行消化作用的溶酶体。
在每个植物细胞内,液泡的大小、形状和数目相差很大,甚至其中的颜色也不相同。在幼龄细胞中,液泡的体积很小,用光学显微镜很难发现。随着细胞的生长,这些小液泡逐渐增大和合并,在成熟的细胞中,通常只有一个大的中央液泡,它可达细胞体积的
90%
。液泡是由一层单位膜包围的细胞器,这层膜叫液泡膜,它使液泡的内含物与细胞质分隔开。液泡膜虽然在形态上与细胞膜无区别,但它们的通透性和物理性质是有区别的。液泡膜含不饱和脂肪酸的比例较高,它所含的磷脂分子以磷脂酰胆碱为主。
|
| |
|
|
|
|
|