|
DNA的复制
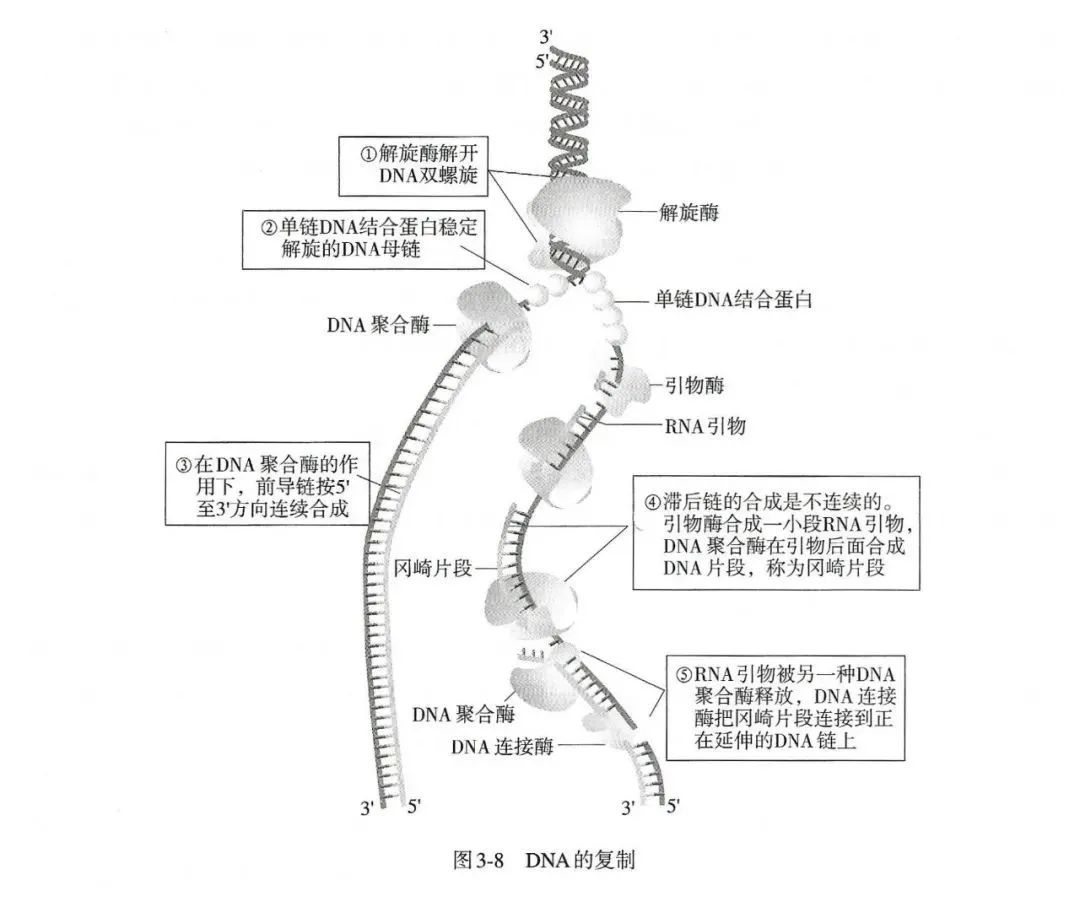
DNA复制最主要的特点是半保留复制、半不连续复制(图3-8)。在复制过程中,原来双螺旋的两条链并没有被破坏,它们分成单独的链,每一条旧链作为模板再合成一条新链,这样在新合成的两个DNA分子中,一条链是旧的而另外一条链是新的,因此这种复制方式被称为半保留复制。
DNA的两条链是反向平行的,一条是5\'→3\'方向,另一条是3\'→5\'方向。在复制起点处,两条链解开形成复制泡(replication bubble ), DNA向两侧复制形成两个复制叉(replication fork)。随着DNA的不断解旋,两条链变成单链形式,可以作为模板合成新的互补链。但是,生物细胞内所有的DNA聚合酶都只能催化5\'→3\'延伸。因此,以3\'→5\'的链为模板链时,DNA聚合酶可以沿5\'→3\'的方向合成互补的新链,这条链称为前导链(leading strand )。当以另一条链为模板时则不能连续合成新链,这条链称为滞后链(lagging strand )。这时,DNA聚合酶从复制叉的位置开始向远离复制叉的方向合成1〜2 kb的新链片段,待复制叉向前移动相应的距离后,又重复这一过程,合成另一个类似大小的新链片段,这些片段被称为冈崎片段(Okazaki fragment)。最后,由另一种DNA聚合酶和DNA连接酶负责把这些冈崎片段之间的RNA引物除去,并把缺口补平,使冈崎片段连成完整的DNA链。这种前导链的连续复制和滞后链的不连续复制在生物细胞中是普遍存在的,称为DNA的半不连续复制。
DNA的复制是一个复杂的过程,需要模板、原料——脱氧核苷三磷酸(dATP、dGTP、 dCTP、dTTP)、酶和蛋白质等多种物质的参与。
DNA复制涉及的第一个问题就是DNA的两条单链要在复制叉位置解开。DNA双链并不会自动解旋,细胞中有一类特殊的蛋白质可以促使DNA在复制叉处打开,这就是解旋酶。解旋酶可以和单链DNA以及ATP结合,利用ATP水解生成ADP时产生的能量沿DNA链向前运动促使DNA双链打开。
解旋酶沿复制叉方向向前推进产生了一段单链区,但是这种单链DNA极不稳定,很快就会重新配对形成双链DNA或被核酸酶降解。在细胞内有大量单链DNA结合蛋白(single strand DNA binding protein, SSB),能很快地和单链DNA结合,防止其重新配对或降解。SSB结合到单链DNA上之后,使DNA呈伸展状态,有利于复制的进行。当新DNA链合成到某一位置时, 该处的SSB便会脱落,脱落的SSB可以重复利用。
(3)DNA拓扑异构酶(DNA topoisomerase)
DNA在细胞内往往以超螺旋状态存在,DNA拓扑异构酶催化同一DNA分子在不同超螺旋状态之间的转变。DNA拓扑异构酶有两类。DNA拓扑异构酶I的作用是暂时切断一条DNA链,形成酶—DNA共价中间物,使超螺旋DNA松弛,再将切断的单链DNA连接起来,不需要任何辅助因子;DNA拓扑异构酶II能将负超螺旋引入DNA分子,该酶能暂时性地切断和重新连接双链DNA,同时需要ATP水解提供能量,如大肠杆菌中的DNA旋转酶(DNAgyrase)。
引发酶在复制起点处合成RNA引物,引发DNA的复制。它与RNA聚合酶的区别在于引发酶可以催化核糖核苷酸和脱氧核糖核苷酸的聚合,而RNA聚合酶只能催化核糖核苷酸的聚合, 其功能是启动DNA转录合成RNA,将遗传信息由DNA传递到RNA。
(5)DNA聚合酶(DNA polymerase)
DNA聚合酶最早是在大肠杆菌中发现的,以后陆续在其他原核生物中找到。它们的共同性质是:以脱氧核苷三磷酸(dNTP)为前体催化DNA合成;需要模板和引物的存在;不能起始合成新的DNA链;催化dNTP加到延伸中的DNA链的3\'-OH末端;催化DNA合成的方向是5\'→3\'。
DNA连接酶是1967年在三个实验室同时发现的。它是一种封闭DNA链上缺口的酶,借助ATP或NAD水解提供的能量催化DNA链的5\'-磷酸基团的末端与另一条DNA链的3\'-0H生成磷酸二酯键。只有两条紧邻的DNA链才能被DNA连接酶催化连接。
所有DNA的复制都是从固定起始点开始的,而目前已知的DNA聚合酶都只能延长已存在的DNA链,而不能从头合成DNA链,那么一个新DNA的复制是怎样开始的呢?研究发现,DNA 复制时,往往先由引发酶在DNA模板上合成一段RNA引物,再由DNA聚合酶从RNA引物3\'端开始合成新的DNA链。对于前导链来说,这一引发过程比较简单,只要有一段RNA引物,DNA聚合酶就能以此为起点一直合成下去。但对于滞后链来说,引发过程就十分复杂,需要多种蛋白质和酶的协同作用,还牵涉冈崎片段的形成和连接。
滞后链的引发过程通常由引发体(primosome)来完成。引发体由6种蛋白质共同组成,只有当引发前体与引发酶组装成引发体后才能发挥其功效。引发体可以沿着滞后链分叉的方向移动,并在滞后链模板上断断续续地引发生成滞后链的引物RNA。由于引发体在滞后链模板上的移动方向与其合成引物的方向相反,所以在滞后链上所合成的RNA引物非常短,长度一般只有3〜5个核苷酸。
在同一种生物体细胞中这些引物都具有相似的序列,表明引发酶要在滞后链模板上比较特定的位置才能合成RNA引物。DNA复制开始处的几个核苷酸最容易出现差错,用RNA引物即使出现差错最后也要被DNA聚合酶I切除,不影响DNA复制的准确性。
RNA引物形成后,由DNA聚合酶III催化,将第一个脱氧核苷酸按碱基互补配对原则加在RNA引物3\'-0H端,而进入DNA链的延伸阶段。
DNA新链的延伸由DNA聚合酶III所催化。为了复制的不断进行,解旋酶须沿着模板前进, 边移动边解开双链。由于DNA的解链,在DNA双链区势必产生正超螺旋,在环状DNA中更为明显,当达到一定程度后就可能造成复制叉难以再继续前进,但在细胞内DNA的复制不会因出现拓扑学问题而停止,因为拓扑异构酶会解决这一问题。
随着引发体合成RNA引物,DNA聚合酶III开始不断地将引物延伸,合成DNA。DNA聚合酶III是一个多亚基复合二聚体,一个单体用于前导链的合成,另一个单体用于滞后链的合成,因此它可以在同一时间分别复制DNA前导链和滞后链。虽然DNA前导链和滞后链复制的方向不同,但如果滞后链模板环绕DNA聚合酶III,并通过DNA聚合酶III,然后再折向未解链的双链DNA的方向,则滞后链的合成可以和前导链的合成在同一方向进行。
当DNA聚合酶III沿着滞后链模板移动时,由特异的引发酶催化合成的RNA引物即可以由DNA聚合酶III所延伸,合成DNA。当合成的DNA链到达前一次合成的冈崎片段的位置时,滞后链模板及刚合成的冈崎片段从DNA聚合酶III上释放出来。由于复制叉继续向前运动,便又产生了一段单链的滞后链模板,它重新环绕DNA聚合酶III,通过DNA聚合酶III开始合成新的滞后链冈崎片段。通过这种机制,前导链的合成不会超过滞后链太多,这样引发体在DNA链上和DNA聚合酶III以同一速度移动。在复制叉附近,形成了以DNA聚合酶III二聚体、引发体和解旋酶构成的类似核糖体大小的以物理方式结合成的复合体,称为DNA复制体。复制体在DNA前导链模板和滞后链模板上移动时便合成了连续的DNA前导链,以及由许多冈崎片段组成的滞后链。当冈崎片段形成后,DNA聚合酶I通过其5\'→3\'外切酶活性切除冈崎片段上的RNA引物,并利用后一个冈崎片段作为引物由5\'→3\'合成DNA填补缺口。最后由DNA连接酶将冈崎片段连接起来,形成完整的DNA滞后链。
|
|