|
引言
种群的空间结构是种群生态学、遗传学和适应性进化等一些早期概念和模型中的关键因素。在过去的十年中,空间结构和动力学的含义已经在种群生物学中被广泛重视。目前,生态学家将更多的精力集中在了模型的研究种,这也间接反映了个体、种群和群落空间分布动态对揭示生态过程的仲要性:生态过程之间存在相互影响,因此其空间结构与种群的出生率、死亡率、竞争捕食等对种群的作用同等重要。因此,在生境快速消失的今天,用模型来研究其对种群或是生态系的影响,就显得尤为重用。
在大尺度上,生态学有三种研究方法:理论、集合种群和景观生态学(图1)。理论生态学家已经研究了一系列数学模型来描述某个区域内个体之间的相互作用以及同质空间内生物体的移动范围,在忽略了环境异质性的理想状态下,描述了种群过程产形成的复杂的空间格局和动态(图1a)。相反,景观生态学则一直强调环境的一致性以及研究区域中物理环境的复杂系和差异性,并在这样的条件背景下再去分析资源的分布、生物体的移动等生态过程(图1c)。因此,理论生态学在很多时候难以用实测数据去验证,而景观生态学则缺少令人信服的理论框架。
第三种方法就是今天要重点介绍的集合种群生态学(图1b),这是一种折中的方法:这里,景观被视为理想化的栖息地斑块(碎片)组成的网络,其中物种是通过迁移来链接的离散的多个本地种群。由于物种对栖息地的选择、生活习性以及迁徙能力的不同,因此这个方法可能对一些物种来说是适用的,但是还有一些物种,其种群分布在空间上是更加连续的。
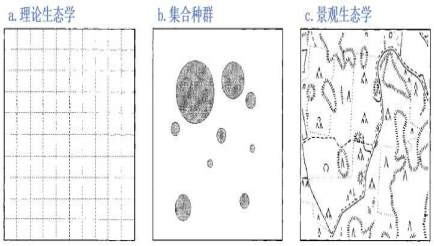
图1三种理论下对环境异质性的假设
“集合种群”一词被广泛应用于任何与种群空间结构相关的研究中,因此“集合种群动态”就包含了所有空间动态。因此,图1的并不能包含所有的与集合种群相关的研究,除了图上展示的这种方法外,近年来还有基于数学模型的、基于保护管理的、基于野外调查或实验的等等。而我个人对集合种群有一个更加严格的定义:即图1中展示的,基于斑块网络假设(patchnetworkassumption)发展起来的所有模型和野外实地研究,尤其是在高度景观破碎化区域的研究。或者我们也可以将此处的集合种群定义为区域内数量不稳定,且生活在离散栖息地斑块的种群的集合。
因此,我们将集合种群称之为种群的种群:就像种群是所有个体的集合一样,集合种群是区域所有种群的集合,那么同样他会在“死亡”(某一个离散的小尺度生境斑块内种群消失,这里我们将其称之为局部灭绝)和“新生”(在新栖息地的种群建立)之间得到平衡。在这样的视角下,集合种群其实很像流行病学:被感染和潜在感染个体就代表已被占据的和可能被占据的栖息地斑块。二者在一些关键过程上的本质是相同的。
集合种群动态,从广义上讲不局限于年龄结构改变、局部灭绝和迁移定居,而是在离散的栖息地斑块中种群的(灭绝和新生)动态。因此这里我们需要从种群的局域灭绝、灭绝-定居动态、斑块面积及破碎化、复杂空间动态、栖息地丧失和破碎化的响应、群落格局以及保护和景观管理等多个角度来理解集合种群。
局域种群灭绝
在集合群中,局域种群灭绝时有发生,并且很多时候会周期性的反复发生,相对应的,在局域灭绝的同时,迁移以及迁移之后在新区域的定居和新种群建立过程也在不断的发生。这一堆灭绝-迁移过程对我们进一步揭示“灭绝”的生态学作用以及理解集合种群动态有着重要的意义。表1总结了在集合种群中可能出现的灭绝过程,并且在以网对庆网蝴蝶(Melitaeacinxia)的研究中,记录到了表里所列出的绝大部分。
顺便说一句,这里说的灭绝分为两种,一种是局域灭绝,也就是集合种群中某一个种群的消失,或者说种群中个体数量变为0;另外一种是整个集合种群的灭绝。前者是集合种群中正常的生态过程,但是如果缺少替代条件或者同时大规模发生,则会直接导致后者的发生。
表1:集合种群灭绝的可能情况
灭绝的尺度
过程的尺度
由随机事件导致的灭绝
由极端事件导致的灭绝
局域尺度
局域尺度
种群数量
环境
基因
栖息地丧失
天敌或竞争者出现
人类活动
局域尺度
集合种群尺度
种群迁移
特定天地出现
集合种群尺度
集合种群尺度
局域灭绝
局域迁移定居
栖息地丧失和破碎化
典型的灭绝事件
影响局域种群数量和环境的随机事件,对整个集合种群的灭绝-迁移过程也会产生影响。如果一个集合种群想要长时间保持稳定或一定的个体数量,那么在区域内需要不断的在新的生境斑块产生新的种群,因为任何一个种群,和个体一样,寿命或者说其存在时间都是有限的。这里同样可以借鉴流行病学的研究来理解:如果一个病毒要长期存在,那么它一定要不断的寻找宿主去生存,因为被寄生的老宿主要么会因为自身抵抗力消灭掉病毒,要么因为病毒而死亡,那么宿主体内的病毒种群同样会在不久后死亡,这里,新的生境就好比一个新的宿主,我们将其称之为,“替代条件”。
替代条件是集合种群长期存在的必要不充分条件,对于一些规模较小的集合种群,所有的种群可能在同一时间灭绝,这就好像毒性极高的病毒,还没来得及扩散就把仅有的宿主全毒死了。另外,在自然界中,并非所有的生境斑块网络中的连通性都是相同的,因此,焦点斑块将决定替代条件是否能够支持集合种群的延续。
对与庆网蝴蝶的研究结果显示,在调查的127个独立的生境斑块网络中,仅有三分之一的网络里斑块数量小于15个,此外,所有的大型的网络中,都出现了随机的灭绝-迁徙事件。而对于小型的生境板块网络而言,集合种群的消失部分是因为这些灭绝-迁徙事件,也有一部分是由于替代条件不成熟。因此,无论替代条件成熟与否,对于较小的生境斑块网络而言,整个集合种群灭绝的概率会更高。
区域性的随机事件会同时影响多个集合中群,从而导致空间上多个种群数量动态的相关性。因此,区域的随机事件减少了种群之间的相独立性,如果随机事件的强度比较大,再加之气候等因素的叠加,很有可能直接使得区域的集合种群难以维系。
此外,着眼于整个集合种群,个体迁出或许是减少种群数量增长率的最大影响因子,若叠加不利的随机环境条件改变,就会增加灭绝的风险,在较小的生境板块中,中因为大量迁出而导致灭绝发生的概率更高。相反,如果附近有一个很大的种群,一直保证一定数量的迁入,那么小种群灭绝的可能就会变得很小。
最近一份关于庆网蝴蝶的研究显示,随着近亲繁殖水平的增加,物种灭绝的风险也在陡增(图2)。之前的研究认为,当某个种群的数量降至很低,而又重新恢复之后,近亲繁殖不太会增加灭绝风险,因为能够在数量瓶颈中恢复,说明他们近亲繁殖所产生的有害等位基因已经被自然选择掉了。然而,庆网蝴蝶的集合种群一直处于“数量瓶颈”中,因为他们单个种群的规模都很小,而目前对于随机遗传和高度破碎化种群的直接和理论都还不足以解释其集合种群中的机制。
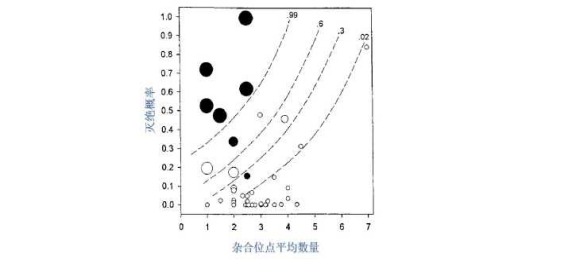
图2灭绝概率与杂合位点数量的关系
研究种群灭绝的最重要的方法之一是有机会从机制上更好地理解栖息地丧失和碎片化单来的影响,二者也是全球生物多样性最大的威胁因子。我们将在下面简要回顾灭绝-迁移动过程之后,再来讨论二者是如何威胁生物多样性的。
灭绝-迁移动态
集合种群是由多个容易灭绝的局域种群组成的,因此其内部保持着局域灭绝和新种群形成的动态平衡。从长时间尺度来看,集合种群内部的动态过程本事是局域种群动态的异步性所维持的,即集合种群中各种群的内禀增长率的差异增加,使得集合种群灭绝的风险随之减少。这种观点与传统的生态学中对单一种群数量稳定的理解相去甚远。对单一种群的研究一般会结合生命表(lifetable)结合关键因子分析,确定与种群密度相关的过程以及种群个体数量稳定的相关和决定性因子。如果这个种群的个体数量急剧减少到某一临界值,或性别比出了问题,那就没得玩儿了。但从集合种群的视角来看,局域灭绝是整个系统数量保持稳定的一个动态过程之一。
已有研究从种群动态、遗传和群落演替等角度探讨了灭绝-迁移动态,而对生态保护而言,最值得我们考虑的一点是集合种群在一个空白的生境斑块网络中的增长率是否是正的,换言之,就是前一节所探讨的替代条件是否成熟。一个小的集合种群的预期增长率既取决于物种的自身属性及其生境的空间结构。而生境的连续性,我们可以通过量化“潜在定居”来计算,但对于集合种群增长率的量化则需要一个考虑更多实际条件的模型来计算。
生物的多样性决定了不同物种对生境要求的特殊性,然而,对特定物种的全面研究还很少,因此相关的学术争论还比较多,这也将是以后研究的一个重点和热点领域。此外,集合种群动态对物种的遗传结构和动态以及物种的适应性进化也具有重要意义。一个比较关键的问题是,在灭绝-迁移过程中,有效遗传种群的大小在多大程度上受到乐影响?此外,基因多样性是否会因为集合种群结构的改变而减小也缺乏足够的实验数据去揭示。
如果种群灭绝或迁移率是由种群的基因特征决定的,那么自然选择可能在局域种群尺度上发生,而在个体层面上则会发生起到相反的效果。还是以庆网蝴蝶举例,这个物种在芬兰有两类宿主植物(婆婆纳属(Veronica)和车前草属(Plantago)),在何种宿主植物上产卵是由基因决定的。因此,对宿主的选择会影响其在生境斑块的选择和建群过程,选择婆婆纳属的个体不会再车前草属的斑块内产卵,反之亦然。于此同时,对宿主的选择也会使得宿主的数量和空间分布产生一定的改变。
生活史特征被认为是由集合群体水平选择决定的,迁移速率的演化就具有了十分重要的意义,因为它直接存在于集合种群动态中。进化生物学家也在研究物种的迁移速率是否会因为栖息地的丧失而加快,这对我们理解生物多样性在变化环境中的保存机制有着重要意义。但是,就目前的生境丧失和环境变化速度来看,进化的速度(种群基因型频率的改变)貌似永远跟不上,很多物种更加倾向于灭绝。
|